BACTRICITY: Bacterial Biosensors with Electrical Output
The metabolically versatile bacterium Shewanella oneidensis adapts to anaerobic environments by transporting electrons to its exterior, reducing a variety of environmental substrates. When grown anaerobically and provided with lactate as a carbon source, S. oneidensis transfers electrons to an electrode of a microbial fuel cell. We sought to engineer S. oneidensis to report variations in environmental conditions through changes in current production. A previous study has shown that S. oneidensis mutants deficient in the mtrB gene produce less current than the wildtype strain, and that current production in these mutants can be restored by the addition of exogenous mtrB. We attempted to control current production in mtrB knockouts by introducing mtrB on lactose, tetracycline, and heat inducible systems. These novel biosensors integrate directly with electrical circuits, paving the way for the development of automated, biological measurement and reporter systems.
Experimental overview
We attempted to develope three inducible systems for electrical current production in S. oneidensis. The first is a chemically inducible system, where LacI or TetR repression of the current-production gene expression in mtrB knock-out (KO) S. oneidensis can be alleviated by the addition of IPTG or anhydrotetracycline, respectively. Our second approach uses temperature-senstive cI which would allow for an increase in electrical output in response to heat. The third is a light-inducible system based on the 2005 UT Austin biological camera.
Using the LacI system, we attempted to interact with engineered S. oneidensis in multiple anaerobic microbial fuel cells which we built, measuring the effects of IPTG on current production in wild type and mtrB KO S. oneidensis containing plasmids expressing inducible or constitutive mtrB.
Results
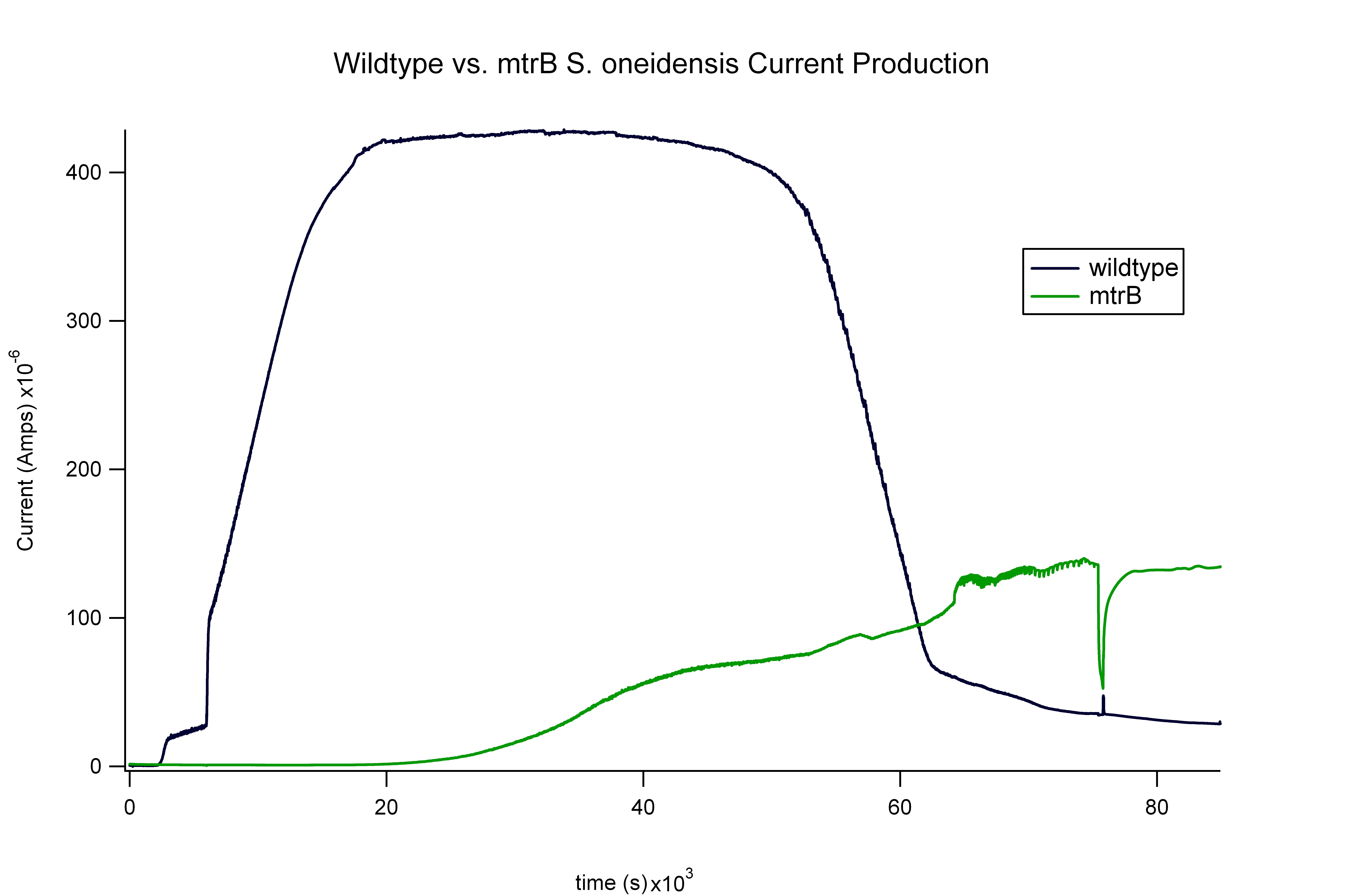
Wildtype vs. mtrB
Co-Culture Experiment
Overview: One possible system for achieving inducible current could be to couple a inducible-lacZ system in E. coli to current production by wildtype S. oneidensis MR1 through the conversion of lactose to lactate. This leaves open the possibility of using a pre-existing E. coli lacZ reporter or creating our own.
In addition to genetically engineering S. oneidensis MR-1 to respond to chemicals and heat, we also sought to take advantage of its natural metabolic pathway of breaking down lactate to produce current. From our wildtype versus mtrB knockout experiments, we found that feeding S. oneidensis MR-1 lactate led to significant current production almost instantly. In wildtype S. oneidensis MR-1, the current production would increase and stay elevated for a period of twelve hours. This result occurred consistently, thus we sought to use controlled lactate release as a way to control current output.
We also knew that wildtype E. coli takes lactose and breaks down lactate. Thus, we hypothesized that it might be possible to couple E. coli’s lactate production with S. oneidensis MR-1’s lactate breakdown to produce current. In addition, a great deal of genetic engineering has already been done on E. coli and the up-regulation or down-regulation of its Lac operon. This means that instead of genetically modifying S. oneidensis MR-1 to respond to chemicals or heat, we can instead take advantage of the vast library of E. coli genetics and couple genetically engineered E. coli with wildtype S. oneidensis MR-1.
In this experiment, we tested different combinations of wildtype E. coli MG1655, S. oneidensis MR-1, and Lac-operon knockout E. coli MC4100 as follows:
1. wt E. coli (MG1655) + wt S. oneidensis (MR1) + lactose
2. wt E. coli (MG1655) + wt S. oneidensis (MR1) + lactate pos. control
3. Lac-operon knockout E. coli (MC4100) + wt S. oneidensis (MR1) + lactose
4. Lac-operon knockout E. coli (MC4100) + wt S. oneidensis (MR1) + lactate pos. control
5. wt S. oneidensis MR-1 + lactose neg. control
6. wt E. coli (MG1655) + lactose neg. control
7. Lac-operon knockout E. coli (MC4100) + lactose neg. control
Based on previous experiments, we would expect current production in combinations 2 and 4 as they both have wildtype S. oneidensis MR-1 and receive lactate. In addition, however, we would expect combination 1 to also produce current. As described above, wt E. coli would break down lactose into lactate, and S. oneidensis MR-1 would break down lactate to produce current.
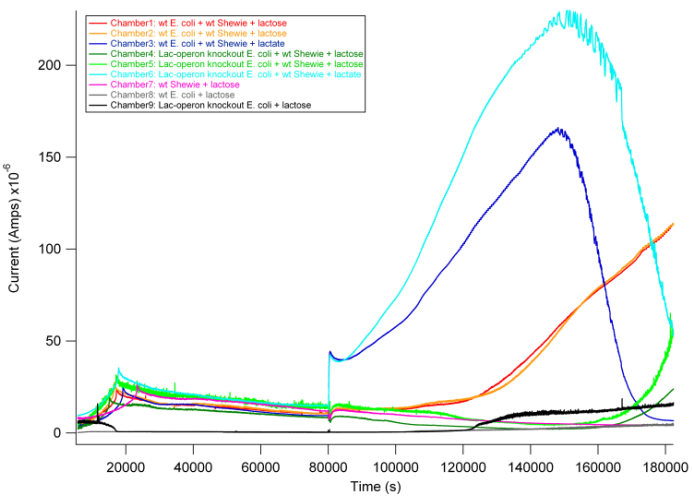
From the data above, we found that combination 1 did indeed produce current with a delay relative to the positive control. The delay can be attributed to the time it takes for E. coli to break down lactose into lactate, thus adding an extra step in the carbon source to current production pathway compared to our positive controls. These results are exciting in that they show a possibility of taking advantage of this cooperative effort to achieve inducible current.
Future directions
Our work with creating a system of inducible electrical output in S. oneidensis has laid the foundations for many different exciting avenues of further inquiry which look to take advantage of a bacteria-computer interface that combines the amazing sensitivity and adaptability of bacteria with the speed and analytical abilities of electricity and computers.
Using the same principles underlying the lac system, the [http://parts.mit.edu/wiki/index.php/University_of_Edinburgh_2006 arsenic biosensor] developed by the University of Edinburgh iGEM 2006 team could be introduced into S. oneidensis, allowing for the coupling of arsenic sensing to an electrical output, a form of a data which is easier to automate and transmit. This could be further extended to other chemical sensing systems, resulting ultimately in an array of different strains S. oneidensis which all respond to the presence of different chemicals with an electrical output that can be monitored by a computer. This could theoretically allow for the remote sensing and analysis of the chemical composition of an environment over time.
Another interesting direction would be the linking of the light-sensing system developed by the UT Austin iGEM team with electrical output in S. oneidensis. In response to variations in light, the amount of electricity produced by S. oneidensis would change. This would allow for the intriguing possibility of not only S. oneidensis conveying information to the computer, but also the computer responding to the S. oneidensis. A simple example would be that in response to a chemical input, S. oneidensis may increase its electrical output. Sensing this increase, the computer could turn on or off a light directed at the S. oneidensis, modifying S. oneidensis's output, creating interesting feedback loops. This could ultimately be developed into more complex communications systems between bacteria and computers. We tried constructing this system over summer, but as the process requires making an EnvZ knockout strain of S. oneidensis, we could not finish it. We did, however, make a few parts to facilitate future attempts.
The possibilities are further broadened by our observations of co-cultures of E. coli and S. oneidensis. Either of the systems described above could be pursued through an alternative alternative strategy of co-cultures. For instance, an array of E. coli which respond to different chemicals by breaking down lactose into lactate could be cultured with S. oneidensis. In response to an increase in lactate, S. oneidensis would begin to produce higher levels of electricity. Co-cultures could also allow for more complex bacteria-computer interactions. This strategy could enable the coupling of almost any E. coli ability to electrical output.
These future directions in which our research can be taken demonstrate some of the exciting possibilities of BACTRICITY!
|  "
"

