|
Home
The Team
Project Report
Parts
Modeling
Notebook
Safety
CoLABoration
|
_meetings
- April 24th 2008
Recruitment of team
- May 8th 2008
attendees
Philipp Mappes, Wolfgang Schamel, Kilian Bartholomé, Katja Arndt, Kristian Müller, Dario Hermida Aponte, Normann Kilb, Robert Gawlik, Simone Weber, Veronika Götz, Kristina Brückner, Kathrin Pieper, Michael Kneib, Moritz Busacker, Sabine Jägle
How to organize the project?
The DNA origamis shall be tested on the established T-cell system. Then we will try to establish an artificial system. Extracellular fragments could be the EGFR or Fas for example. Since the Fas receptor is involved in apoptosis, signaltransduction could be detectable by cell death. Further systems for detecting signaltransduction could be NF-kappaB, the MAP-Kinase-pathway, the Jun-Fos-system or splitenzymes.
DNA-origami structure
How expensive will the synthesis of the NIP coupled Oligos be? Phillip and Michael will contact the company and ask for an calculation of costs. NIP can be ordered by the company Biosearch technology.
Controls for the DNA origamis
- We should produce a DNA origami without NIP-coupled Oligos to check if there also a cell binding occurs (negative control)
- To check the calculated distances between the NIP's on the origami, DNA-origami with fluorophores could be made. By using FRET the distance between the oligo's can be estimated.
- May 23th 2008
- Jun 2th 2008
attendees
Sabine Jägle, Normann Kilb, Robert Gawlik, Kristian Müller, Philipp Mappes
We have talked about...
- To ensure an optimal NIP-scFV-Anti-NIP binding a longer NIP-Linker should be used
- Calculated distances between the NIP's are 6-7 nm.
- Integration of the TCR in the model to achieve a comparison of the sizes
- Searching a suitable transmembraneregion for the artificial receptor
- Calculation of costs for 10 Oligos wirh NIP coupled to the 5' or 3'-end: 3000,-US$ at uncertain coupling efficiency
- => Order presumably from PURIMEX
- Jun 13th 2008
attendees
Michael Kneib, Robert Gawlik, Philipp Mappes, Simone Weber, Kristina Brückner, Sabine Jägle, Kathrin Pieper
Ordering of NIPs
Assembly of the DNA origami with coupled NIPs was discussed and decided:
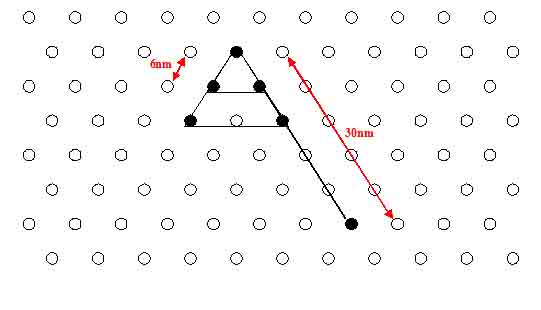
Question: What are the exact distances between the DNA-helices/nod-points? --> Rothemund
The next steps
- searching for literature on TCR-Clustering
- searching for literature on transmembrane-helices, signal sequences
- planning of the artificial system with antibodyfragment, transmembraneregion, splitenzymes, enzyme assay
- EGFR as further test-system
- Jun 16th 2008 10.00h
Preparation of intern presentation on Tuesday
- Jun 17th 2008 12.00h
We have talked about...
- Until Friday we will order the NIP-coupled-Oligos. Consultation of Mahima: Philipp
- We should organize to get the vectors for cloning (Dinah/Christina: Vector with CMV-Promoter) --> Kathrin
- Reproduce Schamels experiment with the TCR --> appointment with Mahima
- Think about a cloning strategy using the BioBrick pre- and suffix
- Literature transmembranhelices WF DeGrado
- Programme for cloning: Emboss (emboss.org)
- Jun 23rd 2008 12.00h
attendees
Robert Gawlik, Philipp Mappes,
Michael Kneib, Kathrin Pieper, Sabine Jägle, Kristina Brückner, Simone Weber
We have talked about...
·
Oligos are ordered. They should be delivered until the 10th of July.
·
Transmembraneregion:
o
Literature about the transmembraneregion
o
Which transmembraneregion can be used? For example EGF-R
o
Signalpeptide?!
o
Paper: Erythropoitetin Receptor Activation by a Ligand-Induced
Conformation Change
·
synthetic transmembraneregion:
o
Which parts shall build the synthetic receptor?
·
Vectors:
o
Genemaps for the Vectors
o
Available Vectors have to be compared to the iGEM-Systeme(Freiburg2007-WIKI)
o
possibly searching suitable vectors
jobs
·
Literature transmembraneregion/signalepeptide : Simone, Kristina,
Sabine
·
Vectorsequences : Philipp, Kathrin,
Michael
·
Registry on the official WIKI:Robert
- July 3rd 12.00h
attendees
Robert Gawlik, Michael Kneib, Kathrin Pieper, Simone Weber, Daniel Hautzinger, Sabine Jägle, Philipp Mappes, Kristian Müller
1. Vectors
Philipp and Kathrin have worked with GCG. In doing so they saw that the fab fragment has iGEM restriction side. The vector from Christina, which we wanted to use, has also 7 iGEM restrictions sides.
We could mutagenesis or gensynthesis to get the iGEM restrictions sides. Genart is offering a special price for gen synthesis. If we want to use this special offer, we have to check which vector they use. It is important that their vector has not any of the iGEM restrictions sides (EcoRI, XbaI, SpeI, PstI) and non of ours restriction sides(NgoMIV, AgeI).
All of the sequences from Mahima arrived by now and they can be checked. A vector wiith CMV-promotor could be also saerched on the wiki 2007.
2. Konzentration der DNA Struktur
theoretically possible: 20 nM, maybe even 200 nM; according we need at least 2 µM to distinct between 2, 3 or 4 NIPs
3. DNA-Origami
- binding measurement: Origami with 4 NIP molecules and 2 fluorophores
- negative control: DNA-Origami without NIP but with fluorophores
- Michael, Simone and Daniel: preparation of M13 phage DNA and assessment of oligo-pool
4. proof of principle: T-cell-receptor activation
- Ca-measurement at 530 nm
- GFP-marker: detection of product after 12 to 18 hours; maybe we can measure FACS at the universities hospital?
5. confocal microscopy
- the MPI requires the absolvation of a training, we don´t have time for that;
- wavelength 488nm and 568nm might be measured at another lab
6. "in general"
- upload more data onto the intern wiki
- meet more frequently
- protocol/write labjournal on wiki
- precise documentation of every generated sample
July 10th, 2008
Attendees:
Simone Weber, Sabine Jägle, Kathrin Pieper, Phillip Mappes,
Norman Kilb, Robert Gawlik, Daniel Hautzinger, Michael Kneib, Kristian Müller,
Kristina Brückner
1. Security instruction
> emergency call: 2000
2.actual state of affairs
- M13mp18 phage DNA was preped
- The Oligios we ordered arrived --> A poll was made, the pool contains all the Oligos except of the 10 which are to be marked with NIP or fluorophor
- Ca2+ - Daniels experiment (making DNA-Origami 1:20 has been reproduced, but we have to check on the AFM if the Origami formed right -> AFM measurement is tomorrow.
- We have to reorder some of the Oligo
3. We need to solve the Origami-concentration problem
- Ca2+We could measure the calcium influx with a flourescence microscope. Therefore we have to find out which dye to use
4. Think about the synthetic receptor system
- we could use a vector which already has the beta-chain of the TCR coupled to the NIP fab fragment and fuse half of β-Lactamase to each of it
- if the receptors get clustered by the NIP the β-Lactamase would be functional
5. substrates for the β-Lactamase
- have to be ordered
- Norman will get some information about diffrent β-Lactamase and order it
6. Transfection Protocol
- available in Kristians lab
7. Positive control for the transfections
- Norman will read about it
8. Searching other vectores
9. second system for multimerisation with different antibodieszweitesn
- Antifluorescin-Antikalin (Arne Skerra) -->
Publication in literature!
- Andreas Blukthuhn (?) --> Dapin (> Kristian will ask him)
10. intracellular domain of EGF-R
- http://www.ebi.ac.uk
--> SRS --> Uniprot
- http://www.expasy.ch
--> swissprot
- Michael will read about it
July 16th 2008 12.00h
Attendees:
Robert Gawlik, Michael Kneib, Kathrin Pieper, Simone Weber, Moritz Busacker, Sabine Jägle, Philipp Mappes, Kristian Müller
1. DNA-Origami:
We reproduced the experiment of Daniel(Origami 1:20). Now we want to test, if we able to form also Origami when we use less Oligos (ratio 1:10, ratio 1:5)
We have to test, if it is possible to use Krebs-Ringer-Hepes buffer for the calcium imaging
We will also produce some Origami coupled with Alexa 488 for first binding measurements. therefore we have also to check were we could measure Alexa 488
Vectors and synthetic receptor system:
still we have not any vector which we can use for the clonings -> we have to check the vector of the Ljubljana-Projekt 2007
Those parts should be planed and ordered:
-Fab-Singlechain
-Which signal sequence coats for the transmembrane domain
-Which transmembran region will we use?
-The transmembrane region could be attached to split-fluorophors, so the receptor clustering could be detected in a fluorescence microscope
- Which linker will we use?
- We should talk to Tillmann Brunner, ZBSA about using the EGFR
July 23rd 2008 12.00h
Attendees
Robert Gawlik, Philipp Mappes, Kathrin Pieper,Sabine Jägle,
Simone Weber.
EGF-Receptor (EGFR):
- The T-cell surface has a lot of big molecules on the surface. Those molecules are much bigger than the receptor itself, therefore they could disturb the interaction between the ligand (e.g. Origami) and the receptor.
-> We should think about using the EGFR as receptor for our clonings, because this one has a relatively big extra cellular domain.
- Not only a part of the EGFR could be used but the whole receptor, which is additionally linked to the NIP fab fragment by a long linker. So the ligand binding domain should be as big as the surface molecules.
- This 3 constructs could be cloned:
1)EGFR + Single chain + one half of beta-Lactamase
2)EGFR + Single chain + second half of beta-Lactamase
3)The whole EGFR fused to the NIP fab fragment. With this construct it might be also possible to activate the receptor by adding EGF. Hence we would have a additionally positive control. But first we would have to find a readout for the activation with EGF.
Fura 2/ Indo-1:
- Both could be used for the Ca2+ signalling. Indo is often used for measurements on the FACS.
- We will use Fura-2-AM for the measurements on the microscope.
Aug. 7th 2008
Attendees:
Robert Gawlik, Philipp Mappes, Normann Kilb, Sabine Jägle,
Kristian Müller, Moritz Busacker.
- problem: Where is the next Ca2+ measurement?
- Take data of the Ca2+-measurement from Prof. Schamel. Titration to get the right concentration?
- pictures from measurement should be handled with "Zeiss Software" (Sabine)
- Carry over T-cells from Mahima (MPI) to kristians lab (Sabine)
- ATG-Biosynthetics synthesizes all parts in a vector without unintentional iGEM restrictionsites
- Invitrogen CCF4AM and CCF2AM for bla detection = LiveBLAzer™
- CCF2AM/CCF4AM 5mg 13.910.00 €
- CCF2AM Loading Kit 200 μg 815.00 €
- LiveBLAzer™-FRET B/G Loading Kit 200μg 690.80€
brochure for further information [http://tools.invitrogen.com/content/sfs/manuals/liveblazer_FRETBGLoadingKit_man.pdf brochure_liveblazer_FRETBGLoadingKit]
Aug. 28th 2008
Sept. 11th 2008
Attendees:
Sabine Jägle, Philipp Mappes, Norman Kilb, Christian Müller,
Wolfgang Schamel, Simone Weber
Themes:
1. CMV
Promotor
Sabine tried to amplificate the CMV Promotor (in the vector) ->
it didn’t work!
Improvements:
- Cut the CMV promoter out (of the vector) and then try again
to amplificate it.
- Prove if we used the right primer.
- Turn the temperature down a bit.
- Use a positive control to be sure the polymerase and buffer
did work
2. Antibodies
Probably we could get some antibodies against NIP from Schamel and his
group, so we could proof if our origamis have the NIP. Normally we
could see the antibodies on the AFM.
3. FACS
We measured the calcium influx on the FACS.
- Unfortunately we didn´t see a calcium influx in
the cytoplasma membrane when we incubated the cells with our origami.
- We have to try to increase the concentration of our origami
- For the next measurement we also have to proof that TA/MgAc
itself has no influence on the calcium influx.
4. Fura-Loading
Because the Fura stain didn´t work when we tried to measure
the calcium influx at the microscope (ZBSA), we wanted to repeat the
staining to see what we should change.
o Again the staining didn´t work ->
We have to ask Nitschke about the conditions.
o We also have to order the Fura-2AM.
5. NIP binding to the
cells
We also repeated the binding measurement at the microscope.
- Like last time both the NIP origami and the control origami
(without NIP) bound to our cells :-(
- We should do the next measurement in Ringer-solution.
Therefore we have to test if the origamis are stable in the
Ringer-Solution with 12,5mM Mg/Ac!!!
- We should repeat this measurement with B-Cells (with a
“NIP-receptor”), to see if the origami (with and
without NIP) bind to those. Maybe some surface structures of the T
cells bind unspecific to our DNA.
-> We can get the B-cells(2558Lδm/mb-1) from Schamel!
Sept. 18th 2008
attendees
Philipp Mappes, Katja Arndt, Kristian Müller, Normann Kilb, Robert Gawlik, Simone Weber, Kathrin Pieper, Sabine Jägle
organisatory
- Bioss agrees to sponsorship
- team picture for wiki
- next meeting: wednesday, Sept. 24th 15.30
- lab: new box-arrangement (Normann)
- table for cloning strategy (Kathrin)
Ca-measurement
- date for measurement at the AFM (Nitschke)
- optionally: measurement at the Henneke lab --> children's clinic (Sabine)
- optionally: measurement at Imaging Facility in the medicin clinic --> searching for authorized contact-persons
Wiki
- carry over the inofficial wiki content to the official one (Robert und Philipp).
- final report about the whole project (everyone)
part-order
- ask for the ATG orders (need to send the plasmid again?) (Philipp)
- obscurities at order BCR-transmembraneregion --> control and correction(Kathrin)
- primer-order to synthesize signalpeptide by PCR (Kathrin)
control of DNA-Origami binding to cell-surface
- add wash-steps after addition of DNA-origami
- cells should keep in Mg2+ for measurement
- possibly the measurement can be carried out by ELISA --> immobilized AK to bind origamis; fluorophors fused to the oligos can be detected or use biotin-fused Oligos to get a detection by streptavidin fused enzyme reaction.
- control: monoclonal AK able to bind DNA-origami (Simone)
Sept. 24th. 2008
-Modeling
-measurement of fura-stained T-and B-cells (ZBSA)
-AFM-measurement at IMTEK?
-consult MPI concerning antibody B1-8
-cloning: CMV+YFP transfection control positive
-order new oligos, this time modified with FluA aswell
Sept. 29th 2008
-forms for traveling on "company" (university) business have to be filled in
-deadlines for short description of the project and standard variance request are close
-can we "digest" (papain/pepsin) the B1-8 IgM pentameres to visualize the antigen pattern on the origami under the AFM?
-received synthesis from GeneArt and ATG:biosynthetics
-FACS still doesn´t show positive results, amount of origami seems still too low<br<
-organization of hotel rooms at special rate
Oct. 7th 2008'
Oct. 17th 2008
attendees:
Philipp Mappes, Katja Arndt, Kristian Müller, Normann Kilb, Robert Gawlik, Michael Kneib
Final report
- structure maybe equal to scientific paper, containing Abstract, Introduction, Materials and Methods, Results
- fusion of all sections we worked in: Origami, AFM, FACS, CA2+/Fura measuremt, cell cultures, modeling, ethics
- quotation of literature through autor and year of publication in the text (Max Muster, 2007), full quotation in references in the end
- deadline Friday Oct. 24th 2008
official wikipediasite
- continue the labjournal on the official page only
- not too much linking: maintenance of the flow of information
- all embedded files should start with "Freiburg2008"
iGEM parts
- uploading the parts starting on Monday Oct. 20th 2008
Oct. 21th 2008
attendees:
Moritz Busacker, Philipp Mappes, Kristian Müller, Normann Kilb, Robert Gawlik, Michael Kneib, Kathrin Pieper, Simone Weber
Cloning:
At least - All the constructes are cloned!!!
construcs:
pMA - signal peptide – scFv-anti-NIP – gggs-Linker – transmembrane region(BCR/EGFR) - X – pMA
X :
- bla1(1/2 beta-Lactamase)-
- bla2 (1/2 beta-Lactamase) –
- split-linker-C-CFP(Cerutan) –
- N-CFP –
- split-linker-C-GFP (split venus)–
- N-GFP –
- luciferase 1 (58) –
- luciferase 2 (57) –
Next steps:
1) We want to test wheather the transmembrane region really is located in the membrane
Therefore we want to clone a construct expressing YFP on the cytoplasmic side of the receptor.
Construct:
- singal peptide – scFv-anti-NIP – gggs-Linker – transmembrane region - split-linker - bla1 - YFP -
2) We also want to test if we able to bring the receptors together using a NIP coupled peptide.
Therefore we need the peptides from Schamels group -> Norman will ask them.
3) Origami-We want to try if the Origami are able to bring two or more receptors together.
Have to produce new Origami -> Michael and Norman
|
 "
"

