|
|

| August |
|
|
WET LAB:
We left PCR of:
- Part 1 of Biopart BBa_I79006
- Biopart cI BBa_C0051
- Biopart 3 (normal) ofBBa_I79006
- Biopart 3 (mut) of BBa_I79006
- Operator of rcnR + RcnA
|
MODELING:
Hill cooperativity 5th Reaction Reminder:
A + B <--> AB
Ka=Keq=[AB]/[A][B]=1/Kd
θ=[AB]/([AB]+[A])=[B]/([B]+Kd)
MWC Model (Cooperativity)
A + nB <--> ABn
Ka=Keq=[ABn]/[A][B]n=1/Kd
θ=[B]n/([B]n+Kd)
log(θ/(1- θ))=nlog(B)-log(kd) …Hill's equation
Suppression mediated by cI:
ρ + nCI <--> ρ:CIn (k+, k-)
Keq=Ka=[ρ:CIn]/[ρ][CI]n
Si ρ0=[ρ]+[ρ:CIn]
… ρ0=[ρ]+Keq[ρ][CI]n
=> ρ= (ρ0/Keq)/((1/keq)+[CI]n)
Flow= k+[ρ][CI]n = K+((ρ0/Keq)/((1/Keq)+[CI]n))[CI]n
Flow= k+([ρ0]/Keq) [CI]n / ((1/Keq)+[CI]n)
=> Vm= k+([ρ0]/Keq) & Kp=1/Keq=Kd
Therefore:
Keq = exp( -ΔG / R T )
k+ = (KB/h) T exp( -ΔG / R T ) = (KB/h) T Keq
Keq= |
2.89517E+17 |
|
KB= |
1.38E-23 |
J/K |
k+= |
1.79764E+30 |
/s |
h= |
6.63E-34 |
J s |
|
R= |
1.9872 |
cal/(K mol) |
ΔG= |
-23810 |
cal/mol |
T= |
298 |
K |
WET LAB:
Gel
We run a gel with the PCR products obtained the day 01-08-08
Purification
From the PCR of the 01-08-08 we took 120 μl to purify DNA in a low fusion point agarose gel in line with the kit.
We run an agarose gel to verify the status of the purified PRC products.

- Molecular Marker
- Part 1 of biopart BBa_I79006
- Biopart cI BBa_C0051
- Biopart 3 (normal) of BBa_I79006
- Biopart 3 (mut) ofBBa_I79006
- Operator of rcnR + RcnA
Restrictions
We left restrictions over night(double and simple) of each biopart.
First Simple Restriction
The bioparts PCR product was cut with 2 simple consecutive restrictions. The reactives and volumes used in the first reaction were the following:
- Part 1; BamH1
Buffer U.... 4 microlts
BSA.......... 4 microlts
BamH1...... 2 microlts
DNA PCR... 15 microlts
H2O......... 15 microlts
Total......... 40 microlts
- Part 2_cI; BamH1
Buffer U.... 4 microlts
BSA.......... 4 microlts
BamH1...... 2 microlts
DNA PCR... 15 microlts
H2O......... 15 microlts
Total......... 40 microlts
- Part 3 Normal; Xba1
Buffer U.... 7 microlts
BSA.......... 7 microlts
Xba1......... 3 microlts
DNA PCR... 28 microlts
H2O......... 25 microlts
Total......... 70 microlts
- Part 3 Mutated; Xba1
Buffer U.... 7 microlts
BSA.......... 7 microlts
Xba1......... 3 microlts
DNA PCR... 28 microlts
H2O......... 25 microlts
Total......... 70 microlts
- Part 4 RcnA; Xba1
Buffer U.... 7 microlts
BSA.......... 7 microlts
Xba1......... 3 microlts
DNA PCR... 28 microlts
H2O......... 25 microlts
Total......... 70 microlts
|
MODELING:
Hill Cooperativity
5th Reaction, conflict
If we consider that:
- Keq = exp (-ΔG / R T)
- k + = (KB / h) T exp (-ΔG / R T) = (KB / h) T Keq
and given that the flow is (k + / Keq) [ρ0] [CI] n / ((1/Keq) + [CI] n), the value of the maximum speed of the flow loses its meaning.
The speed limit is being determined by (k + / Keq) [ρ0], but k + / Keq = (KB / h) * T, and we know that [ρ0] is arbitrary, i.e., Vmax is no longer based on the reaction as such, which does not make sense.
For example: Take the same reaction that we are considering, the maximum speed of the flow of the reaction would be the same with the promoter that has the operators of CI, that if you used one with a random sequence, so, whether we repeated the experiment, with the same temperature and the same concentration of DNA and an equal number of copies of the sequence, the maximum speed reached by the flow would be the same for the real promoter as for for any sequence, without taking any consideration with their affinity for their substrates... That does not makes sense!
The proposed explanation is that the equation used to determine k + does not fit our model. We should explore other possibilities.
WET LAB:
Restrictions
Second simple restriction
Before the second simple restriction we cleaned the product oof the first restriction with the purification Kit.
Due to the purification protocol we knew that the DNA was clean and diluted in 40 μl of buffer, and in order to obtain an efficient restriction we try to dilute the less the DNA-Buffer mix, obtaining the following volumes.
Extraction
Plasmids pRK415 and pBBR1MCS-5 were extracted with the Roche kit(see Techniques).
Cultures
We cultured DH5alfa cells tranfromed with pJet+biopart
|
MODELING:
Hill Cooperativity:
5th Reaction, solving the problem:
The error in the previous approach was that we were considering ΔG to be the same for both equations (for Keq & k+).
We know that the equilibrium depends solely on the difference in the free energy of Gibbs between the substrate and the product (ΔG 'th). The one with less energy will be favored in the balance, while the rate of reaction depends on the activation energy needed for the conversion (ΔG ‡). A reaction reaches equilibrium faster or slower depending on the rate of reaction (depending on the magnitude of its ΔG ‡), but the balance itself does not change.
Thus:
Keq = exp (- ΔG 'º / R T)
k + = (KB / h) T exp (- ΔG ‡ / RT) ≠ (KB / h) T Keq
|
GROUP MEETING
Wet Lab Statusk
Objectives:
- Build the bioparts.
- Transform the bacteria with the construction that we have.
- Design the experiments to test our construction.
- Build the system.
- Collaborate with the modeling group.
To do:
- Extract DNA from the wild type strain to obtain RcnA.
- Get the bioparts catalog.
- Obtain a large amount of plasmid that we can use, and amplify the bioparts.
- Transformation of the bacteria with bioparts.
Currently:
- There are plasmids.
- There are parts already amplified and in a plasmid.
Problems:
- The DNA that we needed was not in the registry.
- The oligos were delayed 2 weeks and a half.
- Issues to extract the plasmid from the colonies.
- Make a PCR ligation with the three parts and amplify with the ends (it did not work).
- With the enzyme used the frequency of spontaneous mutation was increased to about an error every thousand base pairs.
- There is a problem with tetracycline. You get false positives.
What can be done:
- The biopart with RcnA can already be linked to the plasmid.
- For the other construction we will have to link two parts and digest them, then link them with the third part and digest once more, then insert into the final plasmid.
- HindIII can be used with the large biopart to verify the sequence.
Electrodes:
- Will they be specific for Nickel?
|
WET LAB:
Cultures
We left cultures of Biopart 1 in pJEt
Plasmid Extraction
Plasmid RcnA was extracted by alkaline lysis
Gels
We run a 2% Agarose gel with the following samples:
- Molecular Marker (2.5 μl)
- Restriction of part 1_3 (5 μl)
- Double Restriction of RcnA_3 (5 μl) no purified to verify.
- RcnA (purified 5 μl)
- CI (5 μl)
- Part 3 Normal (5 μl)
- Part 3 Mutated (5 μl)
With this gel the parts were verified
PCR
We performed a PCR reaction for RcnA and part 1 using Taq pol.
Transformation y ligation
We took cut RcnA and then it was ligated at vector PBBIMCS_5
2 μl |
vector |
5 μl |
Cut DNA |
4 μl |
buffer |
1 μl |
enzyme (T4 ligase) |
8 μl |
H2O |
20 μl |
Total |
The transformation was performed by the previously mentioned technique and the strain was cultured in two Gentamycine cages(Gm 20)
PCR Cleaning
We cleaned the PCR and it was filled up to 40 μl
|
WET LAB:
4μl of each sample were charged in the following order:
- Molecular Marker
- RcnA 1
- RcnA 3
- RcnA 4
- RcnA 5
- RcnA 6
- Part 1_1
- Part 1_3
- Part 1_6
- Part 1_7
- Part 1_9
<Falta pegar gel segun liber...>
Part 1_1 restriction (Double digestion)
DNA |
36 μl |
EcoR1 |
2 μl |
BamH1 |
2 μl |
BSA |
5 μl |
Buffer |
5 μl |
After double digestion of part 1 we left massive ligation of part 1,2 and 3.
Ligation Recipe:
Part 1 |
5 μl |
Part 2 |
5 μl |
Part 3 |
5 μl |
Buffer 5x |
4 μl |
Ligase |
1 μl |
Total |
20 μl |
Restriction of the other bioparts was performed
RcnA_4 |
|
H2O |
6 μl |
Buffer 2 |
5 μl |
BSA |
5 μl |
DNA |
30 μl |
Xba1 |
2 μl |
HindiIII |
2 μl |
RcnA_6 |
|
H2O |
11 μl |
Buffer 2 |
5 μl |
BSA |
5 μl |
DNA |
25 μl |
Xba1 |
2 μl |
HindiIII |
2 μl |
Part 1_3 |
|
H2O |
6 μl |
Buffer EcoR1 |
5 μl |
BSA |
5 μl |
DNA |
30 μl |
EcoR1 |
2 μl |
BamH1 |
2 μl |
Part 1_6 |
|
H2O |
6 μl |
Buffer EcoR1 |
5 μl |
BSA |
5 μl |
DNA |
30 μl |
EcoR1 |
2 μl |
BamH1 |
2 μl |
Part 1_9 |
|
H2O |
1 μl |
Buffer EcoR1 |
5 μl |
BSA |
5 μl |
DNA |
35 μl |
EcoR1 |
2 μl |
BamH1 |
2 μl |
|
WET LAB:
GEL
2% Agarose gel was run with the product of the massive ligation of the three parts
(we didn't obtain the desired product)
PCR
We pick up a PCR product with the folowing oligos:
a) 1up and 2low
b) 2up and 2low
c) control
The PCR reaction was prepared in the following way:
Reaction 1 |
|
H2O |
10 μl |
Buffer 3.3x |
6 μl |
dNTPs |
4 μl |
Oligo up |
2.5 μl |
Oligo low |
2.5 μl |
Mg (Ac)2 |
3 μl |
DNA |
2 μl |
Reaction 2 |
|
H2O |
9 μl |
Buffer 3.3x |
9 μl |
rTth |
2 μl |
GEL
We run for 1 hr, an 2% Agarose Gel
- Molecular Marker (2.5 μl)
- P1_P2 (oligo 1 up y 2 low) ligation with part 3 (normal) (5μl)
- P2_P3 normal (oligo 2 up y 3 low) (5 μl)
- Negative Control(oligo 1 up y 2 low) (5 μl)
- P1_P2 (oligo 1 up y 2 low) ligation with part 3 (mutated) (5 μl)
- P2_P3 mutated (oligo 2 up y 3 low) (5μl)
- Negative Control (oligo 1 up y 2 low) (5μl)
*The results suggest inspecific primer join.
Cultures
We left cultures of the RcnA + PBBRIMCS_5 ligation
|
WET LAB:
Ligation repeat
Performed in order to obtain at least part 1 and part 2 joined
Part 1 |
5 μl |
Part 2 |
5 μl |
Buffer 5x |
4 μl |
T4DNA Pol |
1 μl |
H2O |
5 μl |
Total |
20 μl |
Ligation PCR of P1_P2
H2O |
33 μl |
Buffer |
5 μl |
dNTPs |
2.5 μl |
Oligo 1 up |
2.5 μl |
Oligo 2 low |
2.5 μl |
Mg(Cl)2 |
2.5 μl |
DNA |
1 μl |
Taq |
|
Total |
50 μl |
*The ligation and the PCR were done ___
Plasmid Extraction
Plasmid PBBIMCS_5 was extracted from the cultures of the last day.
PCR ligation Gel of part1_part2

|
WET LAB:
Ligation part1+PRK415
The tube DR of PRK415 was used along with the following samples of part 1:
- DR Part1_9/pJET (130808)
- DR Part1_6/pJET (130808)
- DR Part1_3/pJET (130808)
Recipe for each ligation tube
Vector |
4 μl |
Part 1 |
8 μl |
Buffer 5x |
4 μl |
Enzyme |
1 μl |
H2O |
3 μl |
Restriction
Linearization of pBB to use it as a control in RcnA+pBB Gel
Recipe for each restriction tube:
H20 |
10 μl |
Buffer |
3 μl |
DNA |
16 μl |
EcoR1 |
1 μl |
|
30 μl |
Plasmid Extraction Gel pBB+RcnA
- Molecular Marker (2 μl)
- [1] Cage2 /11_1
- [2] Cage1 /1_2
- [3] Cage2B /1_1
- [4] Cage2a /10_1
- [5] Cage2b /8_1
- [6] Cage2b /6_1
- [7] Cage1 /5_1
- [8] Cage1 /2_1
- [9] Cage2b /4_1

|
1 |
2 |
3 |
4 |
5 |
6 |
Water |
7 μl |
7 μl |
7 μl |
5 μl |
7 μl |
7 μl |
Buffer |
2 μl |
2 μl |
2 μl |
2 μl |
2 μl |
2 μl |
DNA |
10 μl |
10 μl |
10 μl |
12 μl |
10 μl |
10 μl |
Enzyme |
1 μl |
1 μl |
1 μl |
1 μl |
1 μl |
1 μl |
|
20 μl |
20 μl |
20 μl |
20 μl |
20 μl |
20 μl |
*In order to verify the size of the insert in pBB
Extraction plasmid restrictions (pBB+RcnA) with EcoR1
We used:
- [1] Cage2 /11_1
- [2] Cage1 /1_2
- [3] Cage2B /1_1
- [4] Cage2a /10_1
- [5] Cage2b /8_1
- [9] Cage2b /4_1
Gel
- Molecular Marker (2.5 μl)
- Restriction EcoR1 pBBIMCS-5 2b/4 (2 μl)
- Restriction EcoR1 pBBIMCS-5 2a/10 (2 μl)
- Restriction EcoR1 pBBIMCS-5 2b/5 (2 μl)
- Restriction EcoR1 pBBIMCS-5 2a/14(2 μl)
- Restriction EcoR1 pBBIMCS-5 2a/9(2 μl)
- Restriction EcoR1 pBBIMCS-5 limpio con kit(2 μl)
- Restriction EcoR1 BamH1 PRK415 R1(2 μl)
- Restriction EcoR1 BamH1 PRK415 R2(2 μl
- PCR control1 biopart 1 oligo 1up 2low (5 μl)
- PCR control1 biopart 2 oligo 1up 2low (5 μl)
- Negative Control PCR (5 μl)

Conclusions of the previous gel:
- The Oligos 1up and 2low don't join in unexpected sites
- RcnA wasn't inserted in pBB
- It's necessary to repeat the double restriction of PRK
PCR
- Double ligation
- Control without DNA
- Control without oligos (¿Is there something contaminated?)

We run a gel of this three PCRs and we didn't obtain any product.
Repetición PCR
We repeated the previous PCR reaction using the following recipe:
DNA |
2 μl |
H2O |
32 μl |
dNTP’s |
2.5 μl |
Mg(Cl)2 |
2.5 μl |
Oligo 1up |
2.5 μl |
Oligo 2low |
2.5 μl |
Taq |
1 μl |
Buffer |
5 μl |
|
WET LAB
Restrictions gel pBB+RcnA
- Molecular Marker (2.5 μl)
- [1] Cage2 /11_1
- [2] Cage1 /1_2
- [3] Cage2B /1_1
- [4] Cage2a /10_1
- [5] Cage2b /8_1
- [9] Cage2b /4_
- pBB1
- pBB2
- pBB3

Cultured from a strain result of the transformation
|
We received an e-mail from the Mexico-UNAM-IPN with several questions in regards to our project, here is our reply:
Hello,
We apologize for the late reply, but we had to discuss carefully our answer.
First of all, we think you are confused about what our project really is. We want to make bacteria to modify the extracellular nickel concentration in response to an external signal (AHL in this case), and of course, be able to predict to what extent the concentration of the input signal will affect the amount of nickel in the medium. To achieve this, it is true we have to synchronize our cell population at the beginning. This is easy to do and doesn't represent any technical problems.
We are very conscious of the facts you tell us, first: we know the half-life of the lactones is relatively long (24 hrs as you say). That's why we are including AiiA under a constitutive promoter in our model, which degrades AHL very efficiently. This will ensure AHL does not saturate the medium. Second, we know AiiA does not diffuse freely through the cell membrane. However, we don't need that to happen, as each cell will degrade its own AHL (yes, we are assuming that all AHL will enter a cell within a window of time).
In other words, we do not need to synchronize the bacterial population more than in the first step. We are considering that some cells may respond earlier than others. However, we are assuming that, as we are not changing the physical nor chemical conditions, the proportion of cells responding "earlier" will remain constant, thus allowing us to draw some conclusions of the behaviour of the population as a whole. We hope you see why the synchrony is no longer important for our project.
To summarize what we plan to do, AHL will enter the cell and form a dimer with LuxR (which is under a constitutive promoter, so AHL is the only limiting step). This will start the transcription of cI*, which will repress the expression of RcnA. RcnA is the nickel efflux pump, and thus we are aiming to predict the amount of AHL necessary to get the desired extracellular nickel concentration.
We are doing small moves. At first, we only want to make one successful assay. We hope that in the near future we will be able to use the response time of the system to generate a succession of desired nickel concentrations, thus generating a song.
We hope this letter answers your questions,
LCG-UNAM-Mexico Team
Cuernavaca, Morelos
MODELING:
Reaction 3, AHL:LuxR
Conflict: k3 (ON) <k3 (OFF)?
Reference: Goryachev et al. (2006)
The references they cite for obtaining their parameters were not specific for that kind of parameters, in fact, one mentions the rate of RNA polymerase in HUMAN!
- Check whether the article mentions how they transformed the parameters.
They seem to explain why in their model, the k3(ON) is "so small" to start with:
<<common to all models considered here, is that the stability of the state "off" defined by the constitutive Transcription levels of I and R comes at a price of high value for the critical self Extracellular concentration.>>
This explains the criteria they used to determine their parameters:
<<For each layout we attempted to identify a set of parameters that optimize the functional fitness of the network. The search in the parameter space is constrained by requesting that the kinetic parameters must remain in the biologically realistic range and the resulting network should demonstrate the behavior compatible with our present understanding of the phenomenon quorum sensing.>>
WET LAB
Agarose Gel 1% low fusion point for band purification
- Gel1
- Molecular Weight marker
- [1]
- [2]
- [3]
Ligation PCR parte1+parte2(35 ciclos)
- 1_2 oligo 1up-2lower
- 1_1 oligo 1up-1lower
- 2_2 oligo 2up-2lower
- 1_3 oligo 1up-3lower
- 1_2 oligo 1up-2lower
Recipes:
DNA |
2 μl |
Water |
32 μl |
dNTP’s |
2.5 μl |
Mg(Cl)2 |
2.5 μl |
Oligo 1up |
2.5 μl |
Oligo 2low |
2.5 μl |
Taq |
1 μl |
Buffer |
5 μl |
|
MODELING:
Reaction 4: Natural degradation of cI*
Half life of cI*:
The half-life of a modified cI is of 4 minutes, according to Elowitz & Leibler (2000). They analyze the LAA tail, and JB Andersen et al (1998) conclude that the LAA and LVA tails confer about the same time of life to GFP.
Reaction rate:
Once we get the half-life time of the protein, how do we calculate the rate of reaction and the flow?
The half-life of a reaction (t1/2) is the time it takes for half of the reagents to become products. In a first order reaction, t1/2 is a constant and can be calculated from the rate constant, as follows:
t1/2 =-ln(0.5)/k=0.693/k
This reciprocal relationship between the half life time and the rate constant is very useful when making an estimate of the time a reaction will take to occur. Thus, for k = 0.01/s, the half life time would be about 70 s. For k = 10/s, the half life would be of about 0.07 s or 70 milliseconds. The average life of the reactions of first order is also independent of the initial concentration. If the first half of the molecules react in aprox 20 s, half of the remaining molecules will also take 20 s to react, and so on. The fact that the average lifetime in an unimolecular reaction is a constant means that, at any time of the reaction, a constant fraction of reactive molecules have enough energy to overcome the kinetic barrier and become molecules of product. This makes sense because the energy of a set of molecules is distributed randomly according to a Boltzmann distribution.
* With information from RT Sauer (1999).
NOTE: A first order reaction is the type A → B.
Today we sent an e-mail to Dr. Peter Chivers, an expert in RcnR, to ask him for information in regards to RcnA.
Our mail to Dr. Peter Chivers:
Dr. Peter Chivers,
Hi! My name is Carlos and I'm a student of the Undergraduate Program in Genomic Sciences of the National Autonomous University of Mexico and currently I'm on the fourth year.
The main reason of this e-mail is to get in contact with you, I'll explain you why as briefly as I can. Several students of the program decided to participate in a systems biology competition known as iGEM, and our project focuses on regulating nickel efflux in e. coli. We have to both build the circuit and design a mathematical model to accurately predict the behaviour of our system.
Taking several issues in consideration we decided to regulate RcnA.
Here is the page of our team in case you're interested in reading more about our project:
https://2008.igem.org/Team:LCG-UNAM-Mexico
However we've been having some trouble in finding parameters for our model (such as RcnA's half-life, efficiency, synthesis rate among others). Therefore, having exhausted (or so we believe) the literary resources and having found your name in many of the papers regarding RcnA we decided to contact you. I'll try to explain with more detail the parameters we still lack in case you're interested in providing us with information.
I greatly thank you for taking the time to read this e-mail and await your answer.
Carlos Vargas
Dr. Peter Chivers' reply
Dear Carlos,
Thank you for your e-mail. From what I read on your website, it looks like your team is tackling an interesting project. Unfortunately, there are very few papers on RcnA itself, and none of them have examined the parameters that you need for your model. As far as I know, the groups that have published in the RcnR/RcnA area don't typically do those sorts of measurements so they are unlikely that have the data you need. We have no plans to do these experiments as my lab focuses on the nickel responsive regulators, RcnR and NikR.
I'm sorry that I can't be of more assistance. Good luck with your project, I hope it is well received at the iGEM competition.
Best regards,
Peter
WET LAB
Agarose Gel 1% low fusion point for band purification
- Molecular Weight Marker
- [1] Cage2 /11_1 pBB+RcnA
- [2] Cage1 /1_2 pBB+RcnA
- [5] Cage2b /8_1 pBB+RcnA
- [9] Cage2b /4_1 pBB+RcnA
Gel Purification PCR of pBB+rcnA
- Purification 1
- Purification 2
- Purification 3
- Purification 4 clean PRK
- Positive control
- Negative control
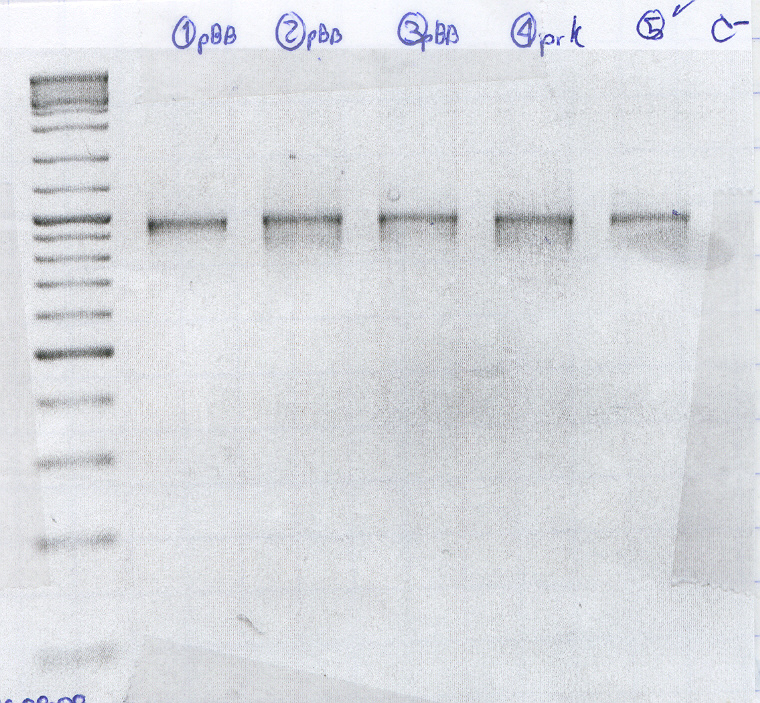
The cells seem to carry the plasmid with RcnA, However one of the negative controls gave product.
All of them were positive, however there are some spurious products.(no photo was added because the low melting gels are quite fragile and we couldn't carry it to the transiluminator with camera.)
Recipe:
DNA |
2 μl |
Water |
32 μl |
dNTP’s |
2.5 μl |
Mg(Cl)2 |
2.5 μl |
Oligo 1up |
2.5 μl |
Oligo 2low |
2.5 μl |
Taq |
1 μl |
Buffer |
5 μl |
A gel with the extraction of the plasmid PRK415+part_1 was run.
Gel Electrophoresis of PCR_Ligation part1+part2
- [5] 1_2 oligo 1up-2lower
- [4] 1_2 oligo 1up-2lower
- [3] 1_2 oligo 1up-2lower
- Molecular marker
- [2] 1_2 oligo 1up-2lower
- [1] 1_2 oligo 1up-2lower

We have got part1+part2 ligation
|
MODELING:
Checking parameters:
Reaction 6
The value that had been found before for k6 (Pl) = 0.20mM / h is in reality the flow of the reaction (which is why the units are mM/h). That is, how many mRNA molecules are being produced per unit of time. The system in which they measured this parameter is a derivative of the plasmid pBR322, pTrc99A. It has the same origin of replication and the number of copies is estimated at 15-20 but under certain conditions where replication is limited it appears to be between 3-5 copies.
Using the previous information we can calculate the value of k+ of the reaction in our system if we consider that each promoter acts independently and we multiply by the ratio between the number of copies of our plasmid and those of their plasmid.
|
MODELING:
To-do List
1. Find Missing Parameters.
- cI Dimerization.
- Nickel efflux . (Estimate?).
- RcnA degradation.
- Nickel Internalization (Experimentally,?).
- Initial concentrations (aiiA, LuxR? (Define arbitrary medium), ? (by number of copies of the plasmid; change to molar), Ni-ext (experimental)).
2. Review tools in SimBiology (sensitivity analysis, parameter estimation, moiety conservation).
3. Stationary states of the system (whether there is multistationarity).
4. Jacobian.
5. Analysis of the stechiometric matrix (also analyze the null space and its transpose).
6. Electrochemical theory (the relationship between electric potential and the concentration of nickel).
7. Electrodes.
->We need to check for advances in the construction of the measurement device we will use.
->Take into account the possibility of asking for support to Dr. Peña.
|
|


|
| |
 "
"

